
El impresionante Tirannosaurus rex, los achaparrados “trilobites” o los misteriosos “ammonites”.
Pero esa visión terminó.
Toda esa curiosidad inocente es destruido por
las garras de la madurez y la ciencia, que inculca, a quien tiene la fuerza y
las ganas, los métodos para investigar, clasificar y estudiar más sobre estos
seres cuyos restos ahora contemplamos y que de niños nos parecieron sencillos y
divertidos.
En este blog paleontológico evaluable y obligatorio,
de carácter científico y divulgativo vamos a ver a algunos de estos seres desde
el punto de vista de todos aquellos que decidieron dar un paso más (o varios)
en el conocimiento de los fósiles.
En esta entrada empezaremos por una parte
realmente interesante y frágil que suele encontrase separada de los muy típicos
caparazones de los ammonoideos. El denominado aptychus.
Se trata de un pequeño órgano hecho de calcita
cuya finalidad no ha estado muy clara. Anteriormente se le habían atribuido siete
propósitos a la utilidad del aptychus: mandíbula inferior, protección de las
gónadas en las hembras, opérculo protector, peso extra, útil para descubrir
presas bentónicas, para el filtrado de microfauna o como medio de propulsión.
En
este artículo se introduciría un octavo propósito, el aptychus funcionaría como
un estabilizador durante el movimiento hacia adelante y hacia atrás en la
natación mediante el pulso a chorro (como la que utilizan los cefalópodos
actuales) evitando el descontrol durante el movimiento.
Introducción
Aptychus o aptychi (sensu stricto) es una valva calcítica, normalmente se
encuentran solas o en parejas, aisladas ( Fig. 1B,
D) o asociadas con ammonites (Fig. 1A,
F), normalmente dentro de la cámara corporal (Fig. 1C,
E). Aptychus ha sido universalmente reconocido como parte integral de los
ammonoideos (Lehmann, 1981, Dagys et al., 1989 y
Morton and Nixon, 1987).
Tienen un aspecto alado y muestran un rango de morfotipos que se pueden ver descritos
e ilustrados por Arkell (1957: L437–L440).

Fig. 1.
En la
siguiente imagen (Fig.
2) pueden comparar algunos morfotipos y sus características, extraídos de Lehmann
(1976: fig. 73).
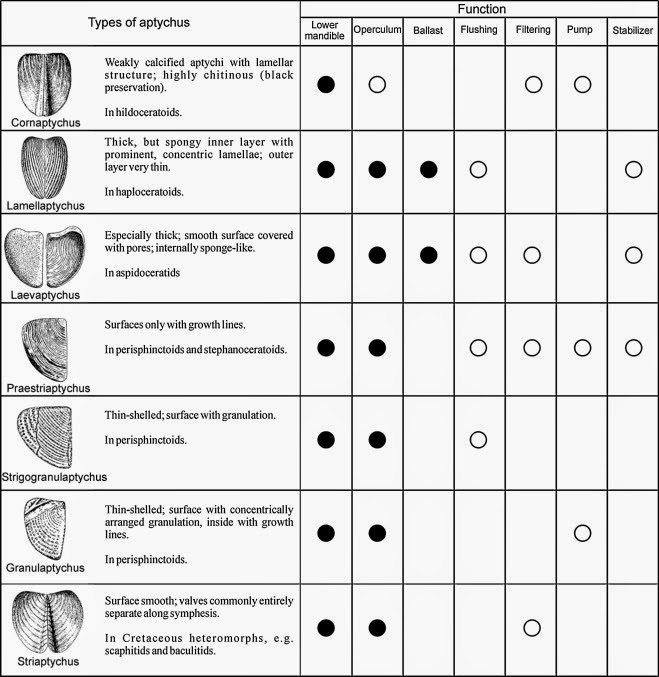
Fig. 2.
Aptychus types characterized by
main features, taxonomic association and proposed functions as explained in the
text. Our assessment of the likelihood that aptychi performed the functions are
as follows: probable (•), possible (o). Illustrations not to scale, modified
from Lehmann (1976: fig. 73) and Trauth (1937: pl. 11: 12).
Funciones atribuidas al aptychus
Describiremos
las diversas hipótesis sobre la función del aptychus en orden histórico.
1. Mandíbula inferior (Meek and Hayden,
1864)
Ésta
función, probablemente derivada de los ancestros de los amontes, debería ser
considerada como la principal función del aptychi (Lehmann, 1971, Lehmann, 1976 and Dzik, 1981). Ésta
hipótesis ha sido aceptada, revirtiendo su anterior rechazo por Trauth
(1927–1938). Las mandíbulas formarían parte de la masa bucal
(Nixon, 1996),
que tambien contendría la radula entre la mandibula superior y la inferior
(e.g., Lehmann, 1967, Lehmann, 1979, Doguzhaeva y Mutvei, 1992 y Klug and Jerjen, 2012; Fig. 3A).
El aptychus actuaría como una tabla más o menos rígida que retendría la presa
para su fragmentación y posterior deglución.
2. Protcción de glándulas nidamentarias y/o
gónadas en hémbras (Keferstein, 1866)
Ésta idea
fue propuesta por Keferstein
(1866) y
aceptada por Favre (1873),
pero fue inmediatamente puesta en duda (Gray, 1873).
La hipótesis de Keferstein fue descartada en las últimas décadas, dada la
ausencia de evidencias cuantitativas en la anatomía de los tejidos blandos de
los ammonoideos, no hay forma obvia para generar argumentos a favor o en contra
de dicha teoría.
3. Opérculo (Trauth, 1927)
El ajuste de muchos aptychi a las cámaras corporales finales de los caparazones en los cuales se encuentran indican que tales aptychis funcionaron como opérculos de cierre de la cámara protegiendo las partes blandas del cuerpo. Un opérculo podría ser necesario para cubrir sólo las partes principales de la sección de la espiral y así proteger el cuerpo blando de ataques de depredadores. Dejando comúnmente espacios abiertos en la zona umbilical de la espiral superpuesta, presumiblemente para la circulación de agua necesaria en la respiración por branquias y para el hyponema (Lehmann, 1990).
Sobre los tipos de aptychi,
tenemos el grueso Laevaptychus de los aspidoceratids y el Lamellaptychus de los
haploceratoids (sujetos de los experimentos descritos en el artículo original: Ammonite aptychi: Functions and role in
propulsion).
Como opérculo,
el aptychus no solo protegería de un acceso directo al cuerpo, también
soportaría el peristoma de la
carga radial aplicada a la periferia por depredadores alargados que tratasen de
quebrar los flancos de la cámara corporal. La capa exterior, laminar en el lado
convexo del aptychus, probablemente sirviera el aptychus para resistir la
tensión bajo compresión radial durante un ataque.
Un
Aptychus grueso, como los de aspidoceratids y haploceratoids, se habría
beneficiado de las mayores fuerzas de bending y buckling que proporcionan las
configuraciones gruesas. Por lo tanto ante el incremento del espesor para la
función como opérculo, la principal adaptación
para el aptychi grueso, fue volverse más poroso.
4. Lastre para reducir la apertura (Gasiorowski,
1960)
El grosor
y el relativo peso del Laevaptychus de los Aspidoceratidae y del Lamellaptychus
de los Haploceratoidea cuando lo proyectan desde la abertura mediante la masa
cefálica, podría reducir la apertura de los brevidomas durante los periodos de búsqueda y alimentación (Fig. 3C). Gasiorowski
(1960: 70) propone que un aptychi grueso actúa de lastre, pero no
describe la proyección de dicho aptychus más alla de la abertura. Esta modificación, que incrementa la importancia del aprychi como lastre, fué sugerida mucho más tarde por Morton and Nixon
(1987). Un aptychi móvil permitiría a una gran variedad de comportamientos (Schweigert, 2009).
5. Caza de presas bentónicas (Lehmann, 1976)
En
algunos ammonites, el aptychus (o anaptychus) pudo haber funcionado como un
hidroplano diseñado para mantener al
ammonite nadando hacia adelante por encima del suelo marino, donde produciría
corrientes que descubrirían a la microfauna en una columna de agua para su
captura (hábitat epidemersal de los amontes de Westermann, 2013)
(Fig. 3C).
Las condiciones estáticas asociadas a esta función son investigadas
experimentalmente y discutidas en el artículo original (Ammonite aptychi: Functions and role in
propulsion).
6. Filtrado de microfauna (Morton and
Nixon, 1987)
El aptychus
podía funcionar un como dispositivo de rigido que crearía una estrecha
hendidura las mandíbulas para el filtrado de microfauna. Esta función fue
apoyada por Kruta et al.
(2011) en Baculites del estudio de los restos de multiples
pequeños organismos preservados en el interior de la masa bucal.
7. Bomba de propulsión a chorro (Westermann, 1990)
El
aptychus podría haber actuado como una bomba para producir la presión necesaria
en el chorro implusor, es decir, una tabla plegable o un paleta con bisagras como alternativa a
la necesidad de mover todo el complejo cefálico como en los Nautilus.
La masa en movimiento dentro de la cámara corporal es mucho menor y compensaría
unos momentos de fuerza más altos, causados por un movimiento más largo debido
a las cámaras corporales de los ammonoideos. La fuerza de este chorro podría
haber provocado una velocidad de natación modesta en algunos ammonites. El aptychi
funcionaría de esta manera, sin embargo, pudo haber producido un flujo casi
continuo de agua para ventilar las branquias, homólogamente a la acción de las
alas-embudo del Nautilus (O’Dor et al.,
1990), en lugar de producir el empuje propulsivo con el
hyponoma.

Fig. 3.
|
Reconstrucción de un
ammonite con aptychus (Ap) grueso actuando en diferentes funciones. Sección
sagital basada en modificaciones de Dzik (1981: fig. 9a). A. Complejo cefálico parcialmente retcostado para
una natación trasera Partially retracted cephalic complex for resting and
backwards swimming. B. complejo
cefálico proyetado con movimiento independiente del aptychus, funcionando de
estabilizador moviendose en armonía con el pulso a chorro para prevenir las
posibles turbulencias, al mismo tiempo que actúa de contrapeso y plano de
dirección para la natación. C.
Complejo cefálico proyectado con aptychus proyectado actuando como bomba de
propulsión y como plano de inmersión durante el movimiento frontal.
-Ct: ctenidia,
-Rm: músculos retractores
-Ra: rádula.
Las flechas indican la dirección del
movimiento.
8. Estabilización ante balanceos (este trabajo)
En este
artículo se propone que algunos aptychi functionaron como estabilizadores
horizontales activos frente a las turbulencias causadas por la propulsión a
chorro (Fig. 3B),
principalmente durante la natación delantera. Se asume que estos ammonites
tenían sistemas de propulsión generados por un chorro de agua a presión que se
generaría en el cuerpo del animal; la tendencia de un ammonite de rotar en
torno al centro de estabilidad de la concha, habría sido más fuerte que en los Nautilus debido a la mayor longitud
en su cámara corporal. Debido a una alimentación a base de animales bentónicos,
probablemente fuese necesaria una orientación del caparazón constante en
relación con el sustrato, el balanceo tuvo que tratar de reducirse al mínimo para
maximizar la eficacia en la caza.
Cuando
era proyectado desde la apertura, el aptychus podría haber proporcionado al
mismo tiempo estabilidad vertical, cuando el ammonites buscase alimento por
encima del lecho marino (2.5 Función de caza de presas bentónicas), como podría
haber tenido las mismas funciones en la natación hacia atrás. Este podría haber
sido otro caso de múltiples funciones de aptychi: lastrado, manejo y
estabilización.
Experimentos sobre la natación frontal
El objetivo de los experimentos
es poner a prueba dos atributos principales de los ammonites planispirales,
especialmente planorbiconic (anteriormente “planulate”; ver Westermann, 1996)
ammonites, como una función de la longitud angular de la cámara corporal:
– Atributo 1. Rotación en torno al centro de
estabilidad del caparazón (balanceo o “eje de carro”) relacionado con varias
funciones indicadas anteriormente. En estos experimentos se asume que el centro
rotacional de la concha se encuentra cerca de los centros de masa y de flotabilidad de la
misma. Esto no sería una suposición irrazonable en la mayoría de los casos (Chamberlain,
1980a). Establecer rotaciones causadas por la variación en la
posición del cuerpo en si, descansando dentro de la cámara corporal calcítica,
a la posición del cuerpo extendido fuera de la misma. Experimentamos las rotaciones en conchas mesodomicas
y brevidomicas. Así como la presencia o no de aptychi de diferentes
grosores: Laevaptychus y Lamellaptychus. Esto nos dirá cuanto “lastre” en el
peso del aptychus se necesita para mover la abertura a una posición cercana al
sutrato y sus efectos sobre las diferentes nataciones.
– Atributo 2. Máximo potencial para la velocidad en
la natacción delantera. El experimento sobre la rotación de la concha puede ser
usado junto a medidas de los momentos de aplicación de la fuerza de chorro,
fuerza de arrastre, flotabilidad, y peso de la concha para estimar la velocidad
de la natación frontal en animales con tamaños de concha similares a los
probados en los experimentos.
1. Términos y parámetros
Variables
y parámetros usados en las imágenes y en el texto:
• B: lastre;
• B1: peso tejido
blando;
• B2: peso aptychus;
• CA: centro de la
espira del caparazón
• CB: centro de
flotabilidad
• CM: centro de masa
• D: diámetro de la concha;
• f: distancia entre CB y CM;
• JT: empuje
producido por chorro;
• OB: ángulo de rotación de apertura;
• OR: ángulo de apertura
•OR1: reposando;
•OR2: mínimo;
•OR3: natación frontal;
• VP: ángulo ventral del
• WT: peso efectivo
• α: ángulo de inclinación de la concha

Fig. 4.
Designs
for the in-air experiments with mesodomic and brevidomic, planorbiconic
Ammonitina with a diameter D = 250 mm, a
volume of 2000 cm3 and tissue mean density of 1.65 g/cm3. The
left half of the batten-shaped bar represents a torsion balance with the pivot
at the buoyancy centre CB, where the apparatus in suspended. Under water
weight, mass WT, is
attached at the mass centre, CM. Torque
is produced by the ballast B attached at the left end of the bar
representing the body protruding from its resting orientation OB1 partially
out of the aperture to OB2. B includes tissues only (B2) or in
addition a variety of Laevaptychus (B1). Their
masses are combined and calibrated to a single mass (B1 and B2). The
restorative moment on f opposes the moment of WT. This
measures the ballast required for lowering the aperture from its resting angle OR1 (shown
for a mesodome at 90° and a brevidome at 60°) to the angle OR2. Forward
swimming by jet thrust JT requires
a low ventral peristome angle VP < 25°
for the recurved hyponome and sufficient stability ST to
counter opposing force produced by JT.
Increasing WT enlarges
the spiral pitch angle (α) and lowers VP. Inset:
detailed view of f1 (mesodome)
andf2 (brevidome).
2. Condiciones experimentales
Debido
a la distribución claramente trimodal de longitudes angulares de las cámaras
corporales de los ammonoideos (Westermann, 1996: fig. 3), los ammonoideos
planispirales pueden agruparse por longitud angular de la cámara corporal (LBC)
en tres tipos de concha: (1) brevidomes,
con cámaras corporales de verticilo medio (180-210 °); (2) mesodomes, con longitudes de espiral de las tres cuartas partes
(250 – 300°); y (3) longidomes, con
cámaras corporales de una espiral o más (350 – 400°). La Longitud de la cámara
corporal determinaría el ángulo de la abertura con respecto a la vertical de
una cáscara vacía o cuando el animal estaba totalmente aislado en la carcasa de
protección.
La
espiral logarítmica amplía el diámetro (D) con cada ciclio, definido por un
factor llamado la tasa de expansión de ciclo (W) por Raup (1967), o por la altura de ciclo
apertural en relación con el diámetro correspondiente (H2D) de Parent et al., 2010 y Parent et al., 2012, i.e.,
la altura del ciclo interno o dorso-ventral. Reducción de LBC y
agrandando W aumenta la distancia relativa entre los centros de masa (CM)
y flotabilidad (CB), es decir, la estabilidad índice ST fD.
El momento producido por la fuerza de chorro debe ir acompañado por el momento
creado por flotabilidad y el peso de la masa; de lo contrario el animal comenzaría
a girar fuera de control.
El
índice de estabilidad ST es aproximadamente 0.03 para mesodomes y
0.05 para brevidomes (Saunders and Shapiro, 1986: fig. 8).
La alta estabilidad de Nautilus (ST ≈ 0.09) es debido a su
extremadamente corta camara corporal y a un W alto, que entre ammonoideos
mesozoicos probablemente sólo es comparable con ancyloceratines excepcionales y
algunos lytoceratines. Las cámaras corporales pequeñas y/o W alto son más
frecuentes en ammonoideos Paleozoicos.
Resultados
Los
resultados experimentales pueden resumirse en la Fig. 5.
Esta figura reune la información de la simulación con un ammonite, planorbiconico,
brevidome e inlado semejante a un
aspidoceratids (D = 250 mm,
volume = 2000 cm3, W = 2,
ST = 0.05). La figura muestra el efecto de orientación
(ángulo de apertura con respecto a la vertical) de la extensión corporal fuera
de la abertura, con y sin aptychus de diferentes pesos. También refleja el
empuje, o fuerza del chorro, que permitiría la longitud del cuerpo sin que se
indujese una excesiva inestabilidad. En todos los casos se mantiene una
flotabilidad neutral (El peso del tejido se reduce en cuanto a un aumento del
peso del aptychus).

Fig.
5.
Stability against
backward rotation by thrust of a hypothetical aspidoceratid with variables and
parameters as in Fig. 4,
withOR1 = 60, OR2 = 5
to 15°, and OR3 = 20
to 40°. Aperture ballast B of soft tissue is augmented with
Laevaptychus varying from 5·103 to 91 × 103 dynes
(up to about 4% of body-chamber volume VBC; shown
by the broken line with circles), which reduces the aperture angle OR3. Various
jet thrusts JT are
then applied countering B for each Laevaptychus ballast
(vertical scales in dynes from the dashed line with circles). The assumed
maximum of the ventral peristome angle (VP = 25°
at OR3 = 29°) required for forward swimming by the
hyponome limits JT.
When the Laevaptychus is moderately thick (45 × 103 dynes,
about 2% of VBC), this
250 mm-diameter hypothetical ammonite can withstand a hypothetical thrust
of 45 × 103 dynes, equivalent to the force producing a modest
forward speed of 20 cm/s (broken diagonal lines), sufficient for foraging.
Conclusiónes
Los
experimentos en mesodomic y brevidomic ammonites cuantifican el
efecto de lastrado de los cuerpos sobresaliendo con un aprychi grueso en:
-(1) una disminución
de la abertura para que el hyponome pueda doblarse hacia atrás bajo la concha
para producir el movimiento horizontal delantero mediante propulsión a chorro.
-(2) proporcionando
estabilidad estática en esta posición de apertura respecto a las fuerzas de
oposición producidas por el chorro.
Las velocidades máximas posibles en la natación frontal para animales subadultos o adultos sin una cámara corporal madura se estiman a partir de estos datos (Fig. 5). Las funciones de filtrado de microfauna, lastre, bomba de propulsión y estabilización contra turbulencias requieren que los aptychus puedan moverse independientemente de la masa bucal.
Algunos
amontes eran capaces de nadar frontalmente, especialmente los Aspidoceratidae y
Haploceratoidea con aptychi grueso y prominente. Además de reducir y
estabilizar la apertura, estos aptychi que sobresaldrían podían haber actuado
como planos de inmersión vertical, estabilización contra las turbulencias
producidas por el impulso a chorro y para filtrado de microfauna en aguas profundas
en mar abierto para su captura. Por lo tanto, un aptychus móvil podría haber
participado en varias funciones, por ejemplo, alimentación, protección y
locomoción.
La
diversidad de los aptychus parecen pequeñas cuando se comparan con la gran
variedad de morfologías en ammonites Mesozoicos, pero parece haber una
asociación consistente entre los tipos de aptychus y los taxones de ammonites. Kruta et al.
(2009) han insistido en que la diferenciación en el desarrollo,
estructura y microestructura de los distintos tipos de aptychus probablemente
indiquen diferencias en el modo de vida y hábitos alimenticios. Estas diferencias van acordes con las
diferencias propuestas en este artículo. These differences are in accordance with the proposed
differences in functions and their combinations discussed above. Su multifuncionalidad
habría limitado la ontogenia y la evolución de aptychi, como se refleja en su
morfología conservadora. Por lo tanto, el aptychi podría ser considerado de
alto valor taxonómico en la sistemática Ammonoidea.
Referencias
C. Burckhardt
Beiträge zur Kenntnis der Jura- und
Kreideformation der Cordillere
Palaeontographica, 50 (1903), pp. 1–144
J.A. Chamberlain Jr.
Hydromechanical design of fossil
cephalopods
M.R. House, J.R. Senior (Eds.), The
Ammonoidea, Academic Press, NY (1980), pp. 289–335
A.S. Dagys, U.
Lehmann, K. Bandel, K. Tanabe, W. Weitschat
The jaw apparati of ectocochleate
cephalopods
Paläontologische Zeitschrift, 63 (1989),
pp. 41–53
L.A. Doguzhaeva, H. Mutvei
Radula of
the Early Cretaceous ammonite Aconeceras (Mollusca: Cephalopoda)
Palaeontographica, A 223 (1992),
pp. 167–177
J. Dzik
Origin of the Cephalopoda
Acta Palaeontologica
Polonica, 26 (1981), pp. 161–191
E. Favre
On some Works relating to a new
Classification of Ammonites
The Annals and Magazine of Natural
History, 11 (1873), pp. 362–374
S.M. Gasiorowski
Remarques sur les Laevaptychi
Annales de la Société Géologique de Pologne, 30 (1960), pp.
59–97
J.E. Gray
Observations on M Favre's paper on a new
classification of ammonites
The Annals and Magazine of Natural
History, 11 (1873), pp. 451–454
W. Keferstein
Cephalopoden. Bronn's Klassen und
Ordnungen der Weichthiere, 3 (2) (1866), pp. 1337–1406 Leipzig-Heidelberg
C. Klug, I. Jerjen
The buccal apparatus with radula of a
ceratitic ammonoid from the German Middle Triassic
Geobios, 45 (2012), pp. 57–65
I. Kruta, I. Rouget, N. Landman, K. Tanabe, F. Cecca
Aptychi microstructure in Late
Cretaceous Ancyloceratina (Ammonoidea)
Lethaia, 42 (2009), pp. 312–321
I. Kruta, N. Landman,
I. Rouget, F. Cecca, P. Tafforeau
The role of ammonites in Mesozoic marine
food web revealed by jaw preservation
Science, 331 (2011), pp. 70–72
U. Lehmann
Jaws,
radula, and crop of Arnioceras (Ammonoidea)
Palaeontology, 14 (1971), pp. 338–341
U. Lehmann
Ammoniten ihr Leben und ihre Umwelt
Stuttgart, Enke-Verlag (1976) 171 pp.
U. Lehmann
The jaws
and radula of the Jurassic ammonite Dactylioceras
Palaeontology, 22 (1979), pp. 265–271
U. Lehmann
Ammonite jaw apparatus and soft parts
M.R. House, J.R. Senior (Eds.), The
Ammonoidea Systematics Association, SpecialVolume 18 (1981), pp. 275–287
U. Lehmann
Ammonoideen
Haeckel Bücherei Volume 2Ferdinand Enke
Verlag, Stuttgart
F.B. Meek, F.V.
Hayden
Paleontology of the upper Missouri
Smithsonian Contributions to Knowledge,
14 (1864), pp. 1–135
N. Morton, M. Nixon
Size and function of ammonite aptychi in
comparison with buccal masses of modern cephalopods
Lethaia, 20 (1987), pp. 231–238
M. Nixon
Morphology of the jaws and radula in
ammonoids
N.H. Landman (Ed.), Ammonoid
palaeobiology. Topics in Geobiology, Vol. 13 (1996), pp. 23–42
R.K. O’Dor, J. Wells,
M.J. Wells
Speed,
jet pressure and oxygen comsumption relationships in free-swimming Nautilus
Journal of Experimental Biology, 154
(1990), pp. 383–396
H. Parent, A.F.
Greco, M. Bejas
Size–shape relationships in the Mesozoic
planispiral ammonites
Acta Palaeontologica Polonica, 55
(2010), pp. 85–98
H. Parent, A.C.
Garrido, G. Schweigert, A. Scherzinger
The Tithonian ammonite fauna and
stratigraphy of Picún Leufú, Southern Neuquén Basin, Argentina
Revue de Paléobiologie, 30 (2011), pp. 45–104
H. Parent, M. Bejas,
A.F. Greco, O. Hammer
Relationships between dimensionless
models of ammonoid shell morphology
Acta Palaeontologica Polonica, 57
(2012), pp. 445–447
D.M. Raup
Geometric analysis of shell coiling:
coiling in Ammonoids
Journal of Paleontology, 41 (1967), pp.
43–65
W.B. Saunders, E.A.
Shapiro
Calculation and simulation of ammonoid
hydrostatics
Paleobiology, 12 (1986), pp. 64–79
G. Schweigert
Die Ammonitenfauna des Nusplinger
Plattenkalks (Ober-Kimmeridgium, Beckeri-Zone, Ulmense-Subzone, Württemberg)
Stuttgarter Beiträge zur Naturkunde,
B267 (1998), pp. 1–61
G. Schweigert
First three-dimensionally preserved in
situ record of an aptychophoran ammonite jaw apparatus in the Jurassic and
discussion of the function of aptychi
Berliner paläobiologische Abhandlungen,
10 (2009), pp. 321–330
A. Steuer
Argentinische Jura-Ablagerungen. Ein Beitrag zur Kenntnis der
Geologie und Paläontologie der argentinischen Anden
Palaeontologische Abhandlungen, 7 (N.F. 3) (1897), pp. 129–222
Aptychenstudien. I-VIII
Annalen des Naturhistorischen Museums in
Wien, 41 (1927–1938), pp. 171–260 42, 121–193; 44, 329–411; 45, 17–136; 47,
127–145
-Westermann, 1990
-Westermann, 1990
G.E.G. Westermann
New developments in ecology of
Jurassic-Cretaceous ammonoids
F. Pallini, F. Cecca, S. Cresta, M. Santantonio (Eds.), Fossili, Evoluzione,
Ambiente (1990), pp. 459–478
G.E.G. Westermann
Ammonoid life and habitat
G.E.G. Westermann
Hydrostatics,
propulsion and life-habits of the Cretaceous ammonoid Baculites
Revue de Paléobiologie, 32 (2013), pp.
249–265
No hay comentarios:
Publicar un comentario